Y190感受态细胞的应用
发布日期:2020/10/18 17:56:53
背景[1-6]
Y190感受态细胞是GAL4系统酵母双杂实验用菌株,MATa型,可直接转化质粒或与MATα型酵母菌株Y187通过mating操作进行蛋白互作验证或筛库试验。Transformation marker为: trp1,leu2,cyh2;报告基因为: lacZ,HIS3,MEL1。Y190-GAL4酵母双杂系统需要两种质粒配套使用:(pGB和pACT2)或(pGBKT7和pGADT7)。
质粒pGB由pGBKT7改造而来,筛选标志为TRP1,用于表达DNA-BD(来自酵母转录因子GAL4N端1~ 174位氨基酸)与目标蛋白(Bait)的融合蛋白;质粒pACT2与pGADT7的结构和功能类似,筛选标志为LEU,用于表达AD(GAL4 C端768~881位氨基酸)与目标蛋白 (Prey)的融合蛋白。
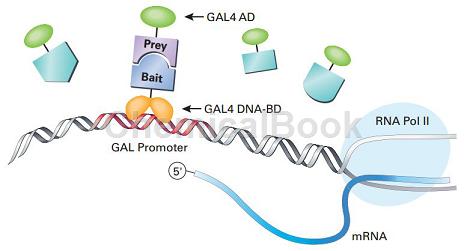
GAL4系统原理:一个完整的酵母转录因子GAL4可分为功能上相互独立的两个结构域:位于N端1~174位氨基酸区段的DNA结合域 (DNA-BD)和位于C端768~881位氨基酸区段的转录激活域(AD)。DNA-BD能够识别GAL4-responsive gene的上游激活序列UAS,并与之结合。而AD可以启动UAS下游的基因进行转录。BD和AD单独存在不能激活转录,但当二者接近时,则呈现完整的GAL4活性,使含有UAS的启动子下游基因转录表达。
正常条件下,BD不与AD结合,将要检测的蛋白质分别与BD和AD融合,形成 bait融合蛋白(bait –BD)和 prey融合蛋白 (prey-AD),如果 bait和 prey发生相互作用,就会促使 BD和 AD的相互接近,形成完整的GAL4,从而激活报告基因的转录。Y190感受态细胞经特殊工艺制作,-80℃可保存三个月,pGADT7质粒检测转化效率>104 cfu/μg DNA。
Y190感受态细胞操作方法:
1. Carrier DNA 在每次使用前要通过加热处理使其变性为单链状态,步骤如下:Carrier DNA 放95℃水浴或金属浴3 min,快速插入冰中,静置3 min,再次放95℃水浴或金属浴3 min,快速插入冰中,静置3 min以上。
2. 取100 μl冰上融化的Y190感受态细胞,依次加入预冷的目的质粒0.5-3 μg,Carrier DNA10 μl,PEG/LiAc 500 μl并吸打几次混匀,30℃水浴30 min (15 min时翻转6-8次混匀)。
3. 将管放42℃水浴15 min (7.5 min时翻转6-8次混匀)。
4. 5000 rpm离心 40 s弃上清,ddH2O 400 μl 重悬,离心 30s弃上清。
5. ddH2O 50 μl重悬,涂板,29℃培养48-96 h。
应用[7][8]
Y190感受态细胞可用于酵母Coronin蛋白与Septin家族成员相互作用研究:
细胞周期(cell cycle)是细胞生命活动的基本过程,其精准调控对生物的生存、繁殖、发育和遗传均具有极其重要的作用。Septin是一个从酵母到人都很保守的基因家族,酿酒酵母中Septin主要组成包括:Cdc3p、Cdc10p、Cdc11p、Cdc12p和Shs1p/Sep7p,Septin结构的变化与细胞周期密切相关。肌动蛋白骨架(actin cytoskeleton)是存在于细胞中复杂的蛋白质纤维网状结构,参与细胞形状的维持、细胞周期、信号转导、增殖分化以及基因表达调控等几乎所有的细胞生理过程。Coronin蛋白(Crn1p)是一种存在于真核生物中的、高度保守的与肌动蛋白骨架调控相关的蛋白质,在肌动蛋白细胞骨架的聚合和解聚过程中都发挥了重要的作用。但目前关于肌动蛋白骨架如何参与细胞周期的调控研究甚少。
本研究以酿酒酵母为研究材料,对酿酒酵母中Coronin蛋白与Cdc3p、Cdc10p、Cdc11p、Cdc12p和Shs1p/Sep7p之间的相互作用进行了初步探讨,并对后续工作提出了设想。主要研究结果如下:
(1)通过酵母双杂交实验筛选出Cdc11p与Crn1p之间具有相互作用。以酿酒酵母S288c基因组DNA为模板,扩增出基因CDC10、CDC11和CDC12的全长序列,大小分别为969 bp,1248 bp和1224 bp,构建酵母双杂交重组质粒p GBKT7-CDC10、p GBKT7-CDC11和p GBKT7-CDC12。p GADT7-CRN1分别同以上三个重组质粒分步转化酵母菌株Y190,并通过对β-半乳糖苷酶活性的检测筛选出Cdc11p与Crn1p具有相互作用。
(2)通过Pull-down实验证实Crn1p与Cdc11p具有相互作用。以p MD19-T-CDC11为模板,扩增出基因CDC11全长序列,构建原核表达重组质粒p ET28a-CDC11。重组质粒p ET28a-CDC11和p GEX-4T-3-CRN1分别转化E.coli BL21(DE3)进行异源表达,得到Cdc11蛋白和Crn1蛋白。最终通过Pull-down实验证实Crn1p与Cdc11p之间的相互作用。
(3)通过免疫共沉淀实验探索Crn1p与Cdc11p之间的相互作用。用限制性内切酶Sac I和Not I酶切重组质粒p ET28a-CDC11和质粒p YES3/CT,构建酿酒酵母表达重组质粒p YES3/CT-CDC11,并转化酿酒酵母菌株BJ5457。同时在CRN1基因的C端融合Myc标签,构建BJ5457 CRN1-Myc且含有p YES3/CT-CDC11的过表达载体的酵母菌株。
通过对其进行诱导前和诱导后形态观察,发现当过表达Cdc11蛋白时,细胞表型异常,细胞子芽延长,且大部分细胞死亡。Co-IP检测时未得到目的条带,且总蛋白条带也较少,猜测由于细胞表型异常导致。因此,本论文初步证实肌动蛋白骨架调控蛋白Coronin与细胞周期调控家族蛋白Septin家族成员Cdc11蛋白具有相互作用,结合前期研究结果:Asr1p蛋白与Coronin蛋白之间具有相互作用,且Asr1p影响细胞周期,我们初步得出一条完整的肌动蛋白骨架调控细胞周期的信号通路。
参考文献
[1]Septin 4, the Drosophila Ortholog of Human CDCrel-1, Accumulates in parkin Mutant Brains and is Functionally Related to the Nedd4 E3 Ubiquitin Ligase[J] . Journal of Molecular Neuroscience . 2012 (1)
[2]Structural and biochemical properties of Sept7, a unique septin required for filament formation[J] . Eldar Zent,Ingrid Vetter,Alfred Wittinghofer. Biological Chemistry . 2011 (8-9)
[3]New insights into the phylogenetic distribution and evolutionary origins of the septins[J] . Ryuichi Nishihama,Masayuki Onishi,John R. Pringle. Biological Chemistry . 2011 (8-9)
[4]Septin structure and function in yeast and beyond[J] . Younghoon Oh,Erfei Bi. Trends in Cell Biology . 2010 (3)
[5]Coronin Switches Roles in Actin Disassembly Depending on the Nucleotide State of Actin[J] . Meghal Gandhi,Vérane Achard,Laurent Blanchoin,Bruce L. Goode. Molecular Cell . 2009 (3)
[6]Actin nucleation and elongation factors: mechanisms and interplay[J] . Melissa A Chesarone,Bruce L Goode. Current Opinion in Cell Biology . 2008 (1)
[7]Nuclear actin dynamics – From form to function[J] . Maria K. Vartiainen. FEBS Letters . 2008 (14)
[8]李国涛. 酵母Coronin蛋白与Septin家族成员相互作用研究[D].云南师范大学,2017.
欢迎您浏览更多关于Y190感受态细胞的相关新闻资讯信息